Ce sont donc des Mammifères, appartenant à l’ordre des Chiroptères, infra classe des Euthériens, classe des Thériens. Les Chiroptères forment un ordre numériquement très important, représenté dans le monde entier, à l’exception des régions polaires et de la Nouvelle-Zélande.Les fossiles trouvés permettent de penser que les chauves-souris dérivent d’une souche insectivore, leur anatomie actuelle nous confirme cette hypothèse. Les Chiroptères sont donc les seuls Mammifères capables de voler activement. Les grands doigts des chauves-souris étendent le patagium et le transforme en ailes cutanées; les muscles moteurs font battre l’aile. Pour obtenir la poussée nécessaire au vol battu, les chauves-souris doivent donc déployer une grande activité et diffèrent profondément des Mammifères planeurs (écureuils volants), qui se contentent d’étaler leur patagium pour profiter passivement de la poussée obtenue par cette surface portante. Chez les planeurs, la force d’inertie joue un rôle essentiel, et en l’absence de courants d’airs ascendants ils perdent continuellement de la hauteur jusqu’à l’atterrissage, alors que les chauves-souris peuvent voler sans limites tant qu’elles ne sont pas épuisées. La forme des ailes varie aussi selon l’espèce : les chauves-souris qui volent rapidement au-dessus des arbres et foncent sur les Insectes, à la manière d’un faucon qui poursuit sa proie en piqué, ont des ailes longues et étroites; celles qui se déplacent lentement entre les buissons, à faible hauteur et dans un secteur restreint, ont des ailes arrondies et larges qui facilitent la recherche tranquille des Insectes posés sur le feuillage. Chez toutes les chauves-souris, la portion antérieure du patagium située devant l’humérus, entre l’épaule, le cou et le poignet, s’incurve vers le bas quand l’aile est étendue et forme un bord d’attaque cintré pour le plan sustentateur (on appelle cette portion cutanée le propatagium).

Chez les Oiseaux, de grandes plumes raides, les rémiges primaires et secondaires, constituent la majeure partie de l’aile, et cependant le propatagium, recouvert de très petites plumes, joue aussi un rôle très important chez eux, sa portion antérieure arrondie représentant également le bord d’attaque. Il est facile de voir ce patagium sur un poulet plumé et qu’on ficelle avant de le mettre à la broche. Il s’arrête à la fin des articulations des membres supérieurs. Il s’avère que cette portion du patagium exerce une fonction essentielle au cours du vol chez les Chiroptères et les Oiseaux. Quelques Oiseaux grands planeurs, tels que les Puffins, ont même à l’articulation du coude un support osseux qui tourne en avant pour favoriser l’extension de la partie médiane du patagium quand l’aile est ouverte.Le squelette des chauves-souris présente une série de modifications en relation avec les facultés voilières. Le radius est l’os principal de l’avant-bras, tandis que le cubitus (ou ulna) n’est plus qu’une fine baguette accolée au précédent et qui contribue cependant à former l’articulation du coude. Les os de la main (métacarpiens) subissent un allongement extraordinaire, sauf le premier. Les phalanges (os des doigts), qui ont subi une réduction numérique, sont également très allongées, mais relativement moins que les métacarpiens. Le deuxième doigt, qui forme le bord d’attaque, ne possède que deux phalanges. Ces os tendent le patagium à la manière des baleines d’un parapluie, mais l’espace qui règne entre les deuxième et troisième doigt est beaucoup plus réduit que les autres. Cette disposition, tout comme celle du propatagium, est destiné à donner un profil correct au bord d’attaque. La plupart des chauves-souris qui font partie du sous-ordre des Mégachiroptères ont sur le deuxième doigt une griffe émoussée qui n’est pas crochue comme celle du premier doigt. Chez quelques espèces, le troisième doigt a lui aussi une griffe.
Les membranes postérieures des chauves-souris ont une structure particulière : la patte subit une rotation de 180 degrés, de sorte que le genoux se plie en arrière et non pas en avant comme chez les autres Mammifères, le pied lui-même étant dirigé vers l’arrière. Quand ce dernier s’appuie sur une surface plane, il est tourné de telle sorte que le talon est plus proche du corps que les griffes des doigts. Une chauve-souris au repos sur une surface plane se tient les ailes pliées et s’appuie sur les poignets et les pieds, tandis que ses genoux sont repliés au-dessus de son dos, comme les genoux d’une sauterelle (qui ne sont d’ailleurs pas leurs homologues). Cette disposition permet l’extension complète du patagium et n’empêche pas les chauves-souris d’être adroites dans leurs mouvements quand elles sont perchées. Elles se montrent en effet très lestes, et si on les dérange, elles courent d’une manière affairée pour se faufiler dans les fissures des grottes où elles dorment. Les espèces frugivores qui passent la journée dans les arbres grimpent d’une façon remarquable et circulent presque aussi aisément que des singes sur leurs branches. Tous les doigts du pied d’une chauve-souris ont une forme et une taille identique; ils portent des griffes acérées avec lesquelles l’animal s’accroche à un support quelconque quand il se repose : les chauves-souris se suspendent à un rameau, à une branche, à une saillie de rocher à l’intérieur d’une grotte, ou bien elles se fixent sur une paroi verticale et le plus souvent se glissent dans une crevasse à l’intérieur d’une caverne ou d’un arbre creux. On a remarqué que les écureuils de la famille des Anomaluridae ont au coude une baguette cartilagineuse qui favorise l’extension du patagium et que les Petauristinae (famille d’écureuils volants d’Australie) ont un processus analogue au poignet. Les chauves-souris possèdent un cartilage similaire au niveau de la cheville : c’est l’éperon, qui forme un angle plus ou moins droit avec la face externe de la cheville et la queue. Ce cartilage acquiert une importance particulière chez les chauves-souris qui n’ont pas de queue ou celles dont la queue est très courte : dans ce cas, en effet, le patagium interfémoral a une surface réduite sur la face interne des pattes, et il s’étend entre l’éperon et l’extrémité de la colonne vertébrale. Sans la présence de l’éperon, ce patagium n’aurait que l’aspect d’une bande de peau très étroite.
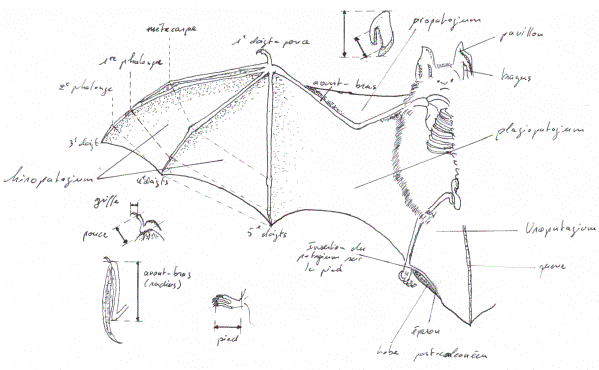

Le squelette zonal et axial est également modifié. La clavicule, os de la ceinture pectorale, est relativement volumineuse et forme une partie de l’articulation de l’épaule; le sternum possède une crête qui augmente la surface d’insertion des muscles utilisés pendant le vol, notamment ceux qui abaissent l’aile et produisent donc une poussée vers le haut. Chez de nombreuses espèces, la fixation de l’aile et des muscles moteurs est encore renforcée par la fusion des côtes supérieures (on retrouve aussi ces caractères chez les Oiseaux). L’anatomie du squelette et des formations épidermiques prouve que les chauves-souris sont les Mammifères les mieux adaptés au vol. Cependant, elles ont conservés les facultés de se mouvoir comme les quadrupèdes, et certaines espèces, les Vampires hématophages (genre Desmodus), sont extrêmement agiles; ces animaux courent sur le corps de leur victime à la manière d’une araignée. L’ordre des Chiroptères peut être considéré comme très dynamique, car ses représentants existent partout dans les régions tropicales et tempérées jusqu’à la limite supérieure des arbres. Les chauves-souris se nourrissent d’Insectes, fruits, fleurs, pollen, nectars, du sang des autres animaux, de la chair des vertébrés terrestres et même de Poissons. Étant donné leur faible taille, elles ont une longévité remarquable. Enfin, elles représentent un plus grand nombre d’espèces que n’importe quel autre groupe mammalien, les rongeurs exceptés.Les Mammifères planeurs utilisent leurs plans sustentateurs pour aller d’un point à un autre, mais aucun ne se nourrit quand il glisse dans l’espace. Il n’en est pas de même pour les Chiroptères, du moins pour les espèces insectivores, c’est-à-dire la majorité (en effet, les frugivores volent pour se déplacer, mais elles se posent pour manger). La plupart des espèces, qu’elles soient carnivores ou végétariennes, cherchent, recueillent et mangent leurs aliments tout en volant. Beaucoup d’espèces insectivores pourvues d’une grande queue recourbent celle-ci en avant de sorte que le patagium interfémoral forme une poche arrondie dans laquelle un gros Insecte peut être manipulé et mis en morceau sans que le vol soit interrompu.Toutes les chauves-souris sont nocturnes et sortent de l’abri où elles ont passé le jour quand survient le crépuscule. Il est mal aisé de trouver une explication à cette habitude. Certes, beaucoup d’Insectes qui forment la seule nourriture des Chiroptères prennent leur vol à la tombée du jour, mais le nombre des Insectes diurnes doit être aussi élevé. La compétition des Oiseaux insectivores serait probablement très importante, si les Chiroptères chassaient de jour; c’est pourquoi il vaut mieux qu’ils concentrent leur activité prédatrice sur des Insectes peu exploités; en effet, mis à part les engoulevents, rares sont les oiseaux insectivores nocturnes. Ce n’est pas dans le dessein d’échapper aux rapaces que les chauves-souris ont adoptés des mœurs nocturnes, car chouettes et hiboux vivent aussi la nuit et en capturent parfois. Certaines chauves-souris américaines gîtent par millions dans des cavernes. Quand elles en sortent, avant la fin du jour, elles sont attaquées par des Oiseaux de proie qui se rassemblent en attendant leur départ. Les Mégachiroptères frugivores et les Microchiroptères mangeurs de pollen et de nectar pourraient aussi facilement trouver leur nourriture le jour que la nuit. Il est vrai que les yeux des chauves-souris sont adaptés à la vision nocturne, à la différence de ceux des oiseaux diurnes; mais est-ce l’adaptation ou l’existence nocturne qui apparut la première ? Ajoutons que tous les Mammifères planeurs sont nocturnes sans exception, et la plupart sont végétariens, sauf les Phalangers volants, qui prennent une certaine quantité de nourriture d’origine animale. Un écureuil volant à la lumière du jour pourrait être une proie facile pour un faucon rapide, et l’abri des ténèbres joue certainement un rôle dans sa survie, mais les acrobaties d’une chauve-souris seraient bien plus déconcertantes pour un prédateur. Pour conclure, il semble probable que les Microchiroptères aient une existence nocturne pour profiter de la masse de nourriture que représentent les Insectes nocturnes. Les autres espèces qui ont un régime différent, les ont exploités secondairement. Il n’y a donc pour l’instant aucune explication plausible de l’existence nocturne des Mégachiroptères. Rien ne prouve ou ne permet de supposer que ces animaux dérivent d’ancêtres insectivores et nous devons simplement nous contenter de cette observation : tous les Mammifères qui volent et planent ont des mœurs nocturnes.Les chauves-souris frugivores ont de grands yeux, et tout permet de croire que leur vision est excellente; à la tombée du jour, elles volent nettement au-dessus des arbres quand elles vont de leur dortoir au gagnage, et leur odorat développé les conduits là où elles peuvent festoyer. Les Microchiroptères ont des problèmes très différents et beaucoup plus difficiles à résoudre pour trouver leur nourriture, étant donné qu’ils chassent dans l’obscurité et près du sol des proies de taille réduite. Ils doivent d’abord localiser, puis capturer les Insectes, tout en évitant de se heurter aux innombrables obstacles qu’ils peuvent trouver sur leur route. Une fois leur faim assouvie, il leur faut s’orienter dans les profondeurs des grottes et autres lieux obscurs qui leur servent de retraite dans la journée. Pendant longtemps, les naturalistes se sont demandés comment les chauves-souris s’orientaient dans les ténèbres, car des yeux mêmes très sensibles ne peuvent suffire dans le noir absolu. Nous savons maintenant que les Chiroptères possèdent d’autres sens qui leur permettent d’avoir une représentation du milieu ambiant aussi détaillée que celle que nous pouvons avoir d’un endroit bien éclairé. C’est un naturaliste italien, Spallanzani, qui, vers la fin du 18ème siècle, fit les premières expériences pour étudier les facultés d’orientation de ces animaux. Dans une pièce remplie d’obstacles, il lâcha des chauves-souris après les avoir aveuglées. Il constata qu’elles volaient aussi aisément que si rien ne leur avait été fait, et notamment qu’elles évitaient parfaitement les embûches. Cela démontrait que les Chiroptères n’utilisent pas leur vue pour s’orienter; toutefois le professeur Spallanzani ne découvrit pas comment ces animaux se dirigeaient; on imagina plusieurs théories sans en apporter des preuves expérimentales. Certains naturalistes pensaient que la surface du patagium, les grandes oreilles et autres formations épidermiques nues, telles les feuilles nasales qui recouvrent le museau de certaines espèces, étaient sensibles aux plus faibles variations de la pression atmosphérique, ce qui aurait permis aux chauves-souris de repérer les objets à distance sans les toucher. Certains poissons possèdent des organes sensoriels qui réagissent aux variations de pression et les rendent capables de percevoir à distance, mais rien de semblable n’a été trouvé chez les chauves-souris.
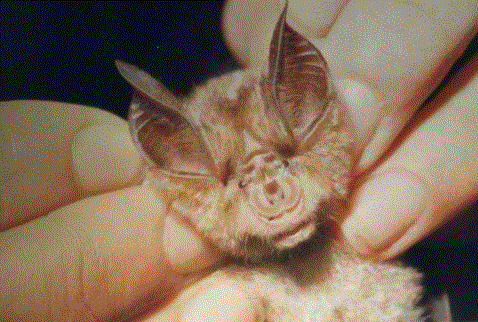
Si l’on approche doucement un objet à quelques centimètres d’une chauve-souris du genre Rhinolophus en train de dormir au plafond d’une grotte, l’animal réagit en fléchissant les genoux, ce qui a pour effet de soulever légèrement son corps (j’ai vérifié ce fait sur un Grand Rhinolophe, et ça marche très bien). Si l’on observait cette réaction à l’approche de la main, on pourrait attribuer ce fait à la perception d’une élévation de température, mais elle se produit aussi quand il s’agit d’un objet inanimé dont la température est identique à celle du milieu ambiant. En outre, la réaction est observée chez des sujets en léthargie, incapables de s’envoler ou même de ramper quand on les dérange de leur perchoir. Ce phénomène peut s’expliquer par le fait que les Rhinolophes émettent les ultrasons par le biais de leur complexe nasale (ensemble de feuilles cutanées dans la région du nez, servant à focaliser les ultrasons); ce qui voudrait dire qu’ils émettent continuellement, d’où la réaction permanente de ce genre à tout objet approchant de son perchoir.Au printemps de 1920, un physiologiste de Cambridge, H. Hartridge, regardait les chauves-souris qui pénétraient dans son appartement par une fenêtre ouverte et venaient chasser les Papillons et d’autres Insectes attirés par les lumières. Il nota que les animaux continuaient à voler quand on éteignait les lampes et qu’ils allaient d’une pièce à l’autre si on laissait la porte entrouverte. Visiblement, il se rendait compte qu’elles ne pouvaient passer sans examiner l’espace d’une façon plus détaillée à l’aide du toucher ou de la vue. Cette observation l’intéressa tellement qu’il entreprit des expériences dans une pièce obscure. Hartridge put constater que ce n’était pas la vue qui leur permettait d’éviter les obstacles. En procédant par élimination, il en vint à conclure que les chauves-souris qui volent dans l’obscurité se rendent probablement compte de la position des objets environnants en émettant des sons ultrasonores qui sont ensuite réfléchis en direction des oreilles. Il publia son hypothèse dans une brève note scientifique, mais comme il avait d’autres recherches en cours, il ne continua pas la vérification de sa théorie, parfaitement exacte, comme nous le savons maintenant. Sa publication n’attira guère l’attention, et pendant vingt ans encore ce sujet de recherche fut abandonné.Quand l’homme inventa l’écholocation pour ses propres besoins, les naturalistes reconnurent que ce système était utilisé par les chauves-souris. Les appareils mis au point par les savants existaient dans la nature depuis des temps très reculés; en effet, les Chiroptères emploient l’écholocation depuis au moins cinquante millions d’années.Le principe de l’écholocation est très simple : on émet un son bref; si les ondes sonores rencontrent un obstacle sur leur trajet, elles sont réfléchies vers la source sonore. Si l’on connaît la vitesse et l’intervalle de temps qui sépare l’émission de la réception, on peut facilement calculer la distance de l’obstacle sur lequel les ondes sonores se sont réfléchies. L’homme a utilisé ce principe pour la première fois en hydrographie, vers 1930. Les sons étaient émis par un appareil placé au fond d’un bateau, et un autre appareil (un hydrophone) recueillait l’écho à son retour du sol sous-marin. Les premiers appareils étaient très primitifs par rapport à ceux de nos jours, et ils n’étaient pas directionnels; le son partait dans toutes les directions et l’écho était réfléchi par l’obstacle le plus proche, c’est-à-dire le fond sous-marin au-dessous du bateau. C’est probablement cette absence d’orientation qui empêcha les naturalistes au courant de cette technique de la mettre en parallèle avec la théorie d’Hartridge. L’écholocation reste simplement une méthode plus rapide que le sondage à main.Peu après 1930, les nécessités de défense aérienne imposèrent la recherche d’un moyen permettant de déceler l’approche des avions ennemis bien avant leur repérage à vue. C’était la seule façon de les intercepter en temps voulu. De plus, il était nécessaire de connaître la position de l’ennemi en distance, direction et altitude. On essaya sans succès des appareils destinés à entendre le bruit des avions, et ces engins mobilisèrent des batteries antiaériennes longtemps après que l’on eut reconnu leur inefficacité. Au cours de la décennie précédente, la télévision avait fait beaucoup de progrès, et la transmission d’images commença quelques années avant la guerre de 1939. Les signaux utilisés en télévision sont transmis par de très brèves impulsions d’ondes électromagnétiques ou ondes radiophoniques, et les ingénieurs électroniciens avaient mis au point des techniques très élaborées pour la production de ces ondes. Les conditions nécessaires à la réalisation d’un appareil d’écholocation étaient donc à peu près réunies, et une équipe de physiciens britanniques dirigée par R.A. Watson Watt eut l’originalité de prévoir les utilisations de cet appareil. Ces hommes eurent suffisamment de persévérance et de volonté pour convaincre les autorités chargées de la défense de l’utilité de leurs idées : ils réussirent à obtenir les crédits nécessaires pour mener à bien leurs travaux. On sait que les ondes radio peuvent passer à travers une certaine épaisseur de matériaux solides, si bien que l’on peut recevoir un programme dans une maison sans avoir à utiliser une antenne extérieur; une partie de ces ondes est réfléchie, notamment par les objets métalliques, et plus courte est la longueur d’onde, plus grande est la proportion d’ondes réfléchies.La radiolocalisation, autrement dit le radar (mot abrégé anglais, RAdio Detecting And Ranging, signifiant détection et télémétrie par radioélectricité), est née de l’application de ses idées.Le principe du radar est le suivant : on émet, par impulsions de très courte durée, un train d’ondes radioélectriques concentrées en un faisceau aussi étroit que possible. Ce faisceau peut être orienté dans n’importe quelle direction, et quand un écho est perçu, l’azimut du faisceau donne le relèvement de l’objet, et la distance combinée à l’élévation du faisceau donne son altitude. On détermine ainsi la position exacte de l’objet dans un espace tridimensionnel. Il est indispensable que le faisceau soit formé d’ondes émises par impulsions séparées, pour que chacune puisse être réfléchie par l’obstacle et produise un écho distinct. La vitesse des ondes radioélectriques (300’000 km/sec) est identique à celle de la lumière, si bien que les impulsions peuvent se succéder à des intervalles très rapprochés, de l’ordre du millième de secondes.Le radar fut tout d’abord utilisé au sol pour détecter la position des avions en vol, mais avec la mise au point d’appareils utilisant des longueurs d’ondes plus courtes, les installations devinrent si petites qu’on équipa les avions avec des radars destinés à fonctionner dans l’espace aérien. Le radar servit donc non seulement à surveiller les autres avions, mais aussi à observer le sol, et il tourna ses regards vers l’enfer que dominaient les bombardiers. Quand les États-Unis d’Amérique entrèrent en guerre, leur potentiel technique et industriel fut orienté sur le perfectionnement du système, et, outre la production d’innombrables appareils, il donna à la radiolocalisation le nom qui l’a fait connaître universellement : le radar.
Pour tous les naturalistes au courant de ce secret de guerre, il était désormais évident que les chauves-souris possédaient très probablement un système d’écholocation identique, à la différence près que les ondes utilisées avaient une vitesse de 330 m/sec et non pas 300’000 km/sec comme les ondes radio. Bien avant la fin de la guerre, les zoologistes américains Griffin et Galambos firent les expériences nécessaires et démontrèrent que la théorie d’Hartridge était parfaitement exacte. Au système d’écholocation qui utilise les ondes sonores, on donna le nom de sonar (mot abrégé anglais, SOund Navigation And Ranging, signifiant détection et télémétrie par ondes sonores).
Le progrès des appareillages électroniques permis l’utilisation pratique du radar et favorisa l’étude du sonar; en effet, sans la technique on ne pouvait guère faire autre chose que constater l’existence d’un système sonar chez les Chiroptères, et il était impossible d’en étudier le mécanisme en détail.
Les chauves-souris aveuglées volent aussi bien que celles dont la vision est normale. En revanche, celles dont on obture les oreilles ou qui ont la bouche muselée sont incapables de voler correctement (sauf pour les Rhinolophes où l’émission des ultrasons se fait par le nez). De toute évidence, ces animaux se guident d’après les échos des sons qu’ils ont émis, l’émission ayant généralement lieu par la bouche, et la réception au niveau des oreilles. Toutefois, ces signaux sonores sont inaudibles pour l’homme, qui doit recourir à des appareils électroniques; ceux-ci les transposent en signaux visibles ou perceptibles; dans les deux cas, on obtient des données analysables que l’on peut enregistrer.Les appareils électroniques ont révélé que les ondes sonores émises par les chauves-souris ont une fréquence très élevée et se trouvent dans la portion ultrasonore de spectre audible, donc au-dessus de l’oreille humaine. Leur fréquence varie entre 30’000 et 100’000 Hertz, et plus; or l’oreille humaine ne peut percevoir que les sons de fréquences comprises entre 16 et 20’000 Hz. En général, elle émet dans une gamme de fréquences assez basse; par exemple, le do du piano fait 256 Hz, et la note la plus haute de l’orchestre, le ré de la petite flûte fait seulement 4’752 Hz. Les chauves-souris ne sont pas les seules à percevoir les ultrasons : d’autres Mammifères peuvent également les entendre. Le sifflet silencieux qui produit des sons inaudibles pour l’homme est utilisé par certaines personnes pour appeler leur chien, et celui-ci l’entend parfaitement. Grâce à l’appareillage électronique, on a appris quantité d’autres choses, comme la longueur de chaque émission. On s’est également aperçu que si le principe du sonar est relativement simple, la réalité vivante présente une grande variété dans les détails non seulement d’une espèce à une autre, mais également d’un individu à un autre. Beaucoup de choses ont été découvertes, mais il y a encore davantage à trouver. Ainsi il est probable que l’on pourra distinguer la plupart des espèces de chauves-souris vivant en Grande-Bretagne (il y en a une quinzaine) ou en France (environ une vingtaine) d’après les caractéristiques de leur sonar; on a même construit un appareil, le détecteur de chauves-souris, qui transpose les ultrasons du sonar en sons perceptibles et permet d’identifier les espèces par leur émission, dont les caractéristiques sont différentes pour chacune. Serait-il possible de déterminer par cette méthode les 900 espèces connues dans le monde ? Nous l’ignorons encore, par manque de données sur les différents sonars.
Dans le radar, les impulsions doivent être espacées pour que l’écho de chacune soit reçu avant le départ de la suivante; cela est essentiel, puisque le principe même du dispositif est basé sur le calcul du temps qui sépare la réception de l’émission. Le sonar de certaines chauves-souris est identique, car au moment où l’animal approche d’un objet, la fréquence des impulsions augmente; au fur et à mesure que la distance diminue, la durée de l’aller-retour des ondes décroît aussi, ce qui rend possible l’augmentation du rythme des impulsions. De cette façon, la chauve-souris reçoit davantage d’informations sur l’objet rencontré.
Comme dans le radar, le flot d’impulsions qui sort par la bouche, ou par le nez, de la chauve-souris est concentré en un faisceau orientable dans plusieurs directions; ainsi l’animal peut examiner l’environnement et reconnaître ce qui peut l’intéresser; les autres Mammifères n’agissent pas autrement quand ils scrutent l’horizon avec leurs yeux.Les premiers ingénieurs radaristes essayèrent de rendre les impulsions aussi compactes que possible; ils voulaient, en effet, leur donner un profil très abrupt au début et à la fin, la variation de zéro au maximum étant presque instantanée; en outre, ils visaient à obtenir des oscillations d’amplitude uniforme à l’intérieur de l’impulsion. Certaines chauves-souris produisent des impulsions voisines de ce type; chez d’autres, en revanche, l’impulsion ne forme pas un bloc uniforme : elle présente des variations d’amplitude et de fréquence, et souvent sa partie terminale vue sur un sonagramme est formée de hachures. Nous ne savons pas si ces irrégularités correspondent à des défauts dans le système sonar des chauves-souris ou à des détails inconnus, mais il semble probable qu’elles augmentent l’efficacité du sonar, qui fournit donc des informations plus précises.
On connaît en détail le sonar d’un très petit nombre d’espèces de chauves-souris appartenant pour la plupart à la famille des Vespertillionidae, largement répandue dans le monde; beaucoup d’espèces communes en Europe et en Amérique du Nord en font partie. Ces animaux émettent par la bouche de brèves impulsions concentrées en faisceau étroit, mais une partie de l’énergie sonore se disperse sur les côtés. Ces impulsions ont une durée de 5 à 10 millisecondes, et dans chacune la fréquence des oscillations varie énormément, puisqu’elle est tantôt de 10 kHz et tantôt de 100 kHz. Les impulsions ont une fréquence modulée, celle-ci étant maximale au début et basse à la fin. Les impulsions se succèdent au rythme de 50 par seconde quand la chauve-souris approche d’un obstacle et sont plus espacées dès qu’elle s’en est éloignée. On a émis l’hypothèse que la modulation de fréquence aide l’animal à distinguer le son émis de l’écho réfléchi, mais il se pourrait aussi que la différence d’amplitude suffise. Selon une théorie ingénieuse, une impulsion à fréquence modulée pourrait interférer avec l’écho de l’impulsion précédemment émise, ce qui produirait des ondes de fréquence moins élevée et de plus grande amplitude; de cette façon, la portée du sonar serait augmentée et le rythme des impulsions pourrait être plus rapide, car elles seraient émises avant que l’écho des précédentes soit reçu. Cela expliquerait l’étonnant précision avec laquelle les chauves-souris évitent les obstacles et capturent leurs proies en plein vol. Toutefois, certaines espèces, quand elles approchent de leur proie, augmentent le rythme des impulsions au fur et à mesure que la portée diminue et raccourcissent la durée de l’impulsion de telle façon qu’elle n’interfère pas avec son propre écho ou un autre; ce fait vient à l’encontre de la théorie précédente. Vraisemblablement, la chauve-souris utilise plusieurs méthodes selon les circonstances, car les impulsions diffèrent par leurs durées et leur fréquence propre, et le rythme d’émission varie également. Les animaux sont vivants, donc beaucoup plus souple que des machines, et ils utilisent au maximum leurs facultés. Ainsi les chauves-souris emploient une longueur d’onde de 20 à 30 kHz quand elles sont en vol de croisière et qu’elles explorent le milieu environnant, car avec ses fréquences l’énergie réfléchie est plus importante. Pour prendre un exemple familier, un marin dont la voix est grave peut aisément héler son bateau ancré au milieu d’une rivière, tandis que la voix aiguë d’une jeune fille aura une portée beaucoup plus faible. Quand une chauve-souris se rapproche d’une proie, elle réduit la longueur d’onde de ses ultrasons, c’est-à-dire qu’elle augmente leur fréquence, et celle-ci atteint 50 à 70 kHz; il lui est alors possible de repérer de tout petits moustiques à une distance de 50 cm et de faire des virevoltes surprenantes pour les capturer. Les chauves-souris rangées dans la famille des Rhinolophidae ont une morphologie bien différente de celle des Vespertillionidae, et par suite leur système sonar est tout autre. Elles sont répandues dans l’Ancien Monde et n’existent pas en Amérique. En Europe, le Grand et le Petit Rhinolophe fer-à-cheval (famille des Rhinolophidae) en sont des représentants caractéristiques. Ces animaux doivent leur nom aux appendices foliacés qui recouvrent le nez. La feuille nasale située au-dessus de la lèvre supérieure a, en effet, une forme plus ou moins régulière de fer-à-cheval. Cet appendice est inséré autour des narines, mais il a des bords libres qui recouvrent la peau garnie de fourrure. La partie supérieure s’amincit en forme de pointe et à sa base existe un lobe, la selle, implanté à angle droit par rapport au plan général de la feuille nasale. La surface et les bords de celle-ci présentent de nombreux replis et des indentations. Ces différents caractères sont propres aux membres de la famille des Rhinolophidae. Dans certaines familles du vaste groupes des Rhinolophidae, il n’y a pas de pointe verticale, mais les circonvolutions de la feuille nasale sont parfois très complexes et l’animal porte un masque des plus étranges.
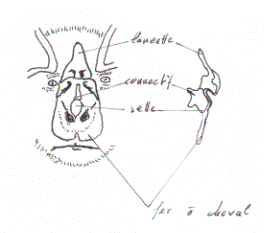
Les chauves-souris de cette famille émettent les impulsions ultrasonores par les narines, et la feuille nasale sert à concentrer ces ultrasons en un faisceau qui est projeté en différentes directions par les mouvements de la tête. Les impulsions sont beaucoup plus longues que celles des Vespertillionidae et durent environ 100 millisecondes, mais leur rythme est plus faible, 4 à 6 impulsions par seconde environ. Elles n’ont pas de fréquence modulée et leur fréquence propre reste constante à environ 80 à 100 kHz selon l’espèce. La longueur des impulsions et leur structure sont telles que ces chauves-souris utilisent l’effet Doppler dans leur écholocation.On a un bon exemple de cet effet Doppler en écoutant le sifflement de la vapeur d’une locomotive qui approche, passe, puis s’éloigne de l’observateur. La fréquence du son émis reste constante, mais sa hauteur semble augmenter au fur et à mesure que la locomotive s’approche, puis descendre une fois qu’elle est passée. Quand elle approche, l’oreille reçoit un grand nombre de vibrations par seconde; le son exact est perçu quand la machine passe devant l’observateur; dès qu’elle s’éloigne, le nombre de vibrations par seconde qui atteignent l’oreille diminue. La fréquence de l’écho reçu par les chauves-souris du groupe des Rhinolophidae semble également augmenter quand l’animal approche d’un obstacle et diminuer quand il s’en éloigne. Il est également possible que les ondes soient produites par l’interférence d’une impulsion émise et de son écho; cela permettrait à la chauve-souris d’obtenir davantage de renseignements sur son entourage. Ce système est aussi efficace que celui des Vespertillionidae et peut-être même supérieur, car un Rhinolophe peut choisir comme perchoir une saillie minuscule sur le plafond d’une grotte, faire une culbute en l’air, s’accrocher à la paroi et replier ses ailes, le tout en une fraction de seconde et dans l’obscurité la plus complète. La production de sons aigus, et notamment d’ultrasons, exige une plus grande quantité d’énergie que la production de sons graves. Le larynx, formé de pièces cartilagineuses, se trouve à la partie supérieure de la trachée-artère. Il est relativement volumineux chez les chauves-souris. Quelques espèces ont un larynx partiellement ou totalement ossifié; par suite, les muscles qui meuvent ses éléments et qui modifient la tension des cordes vocales ont une épaisseur considérable. En outre, l’épiglotte est de forme tubulaire chez les Rhinolophes, et elle s’avance dans les voies respiratoires en arrière de la bouche. Comme nous l’avons vu précédemment, ce genre d’épiglotte n’est pas l’apanage des chauves-souris, puisque celle de nombreux autres mammifères possède la même structure : un chien ne peut pas aboyer tant qu’il n’a pas dégagé son épiglotte et relevé la tête.
Les chauves-souris ne se contentent pas d’émettre des ultrasons; pour communiquer entre elles, elles utilisent aussi des sons de basses fréquences que l’oreille humaine est capable de percevoir. Dans un dortoir de Chiroptères, on entend des cris aigus qui accompagnent les bousculades, de même si on manipule ou dérange ces petits animaux ils protestent en émettant des cris de colère ou de frayeurs. Quand les chauves-souris sont actives et se préparent à prendre leur vol, on peut entendre un très faible bourdonnement. Ce bruit a pour origine un flot d’impulsions se succédant rapidement; chaque impulsion, quoique formée d’une série d’oscillations très brèves, agit sur l’oreille comme une seule onde. Les oscillations qui constituent une impulsion ont une amplitude trop élevée pour que l’oreille de l’homme les perçoive, mais chaque impulsion prise en bloc peut agir aussi comme onde de pression, et c’est la succession rapide de ces ondes qui produit un bourdonnement audible.
Jusqu’à présent, il a été question de l’appareil d’émission du système sonar, mais le dispositif de réception joue aussi un rôle aussi important. Nos connaissances sur ce sujet sont plus réduites. Toutes proportions gardées, les oreilles de la plupart des chauves-souris sont plus grandes que celles des autres Mammifères, sauf exception (les Anglais baptisent le Fennec du nom de “Renard à oreilles de chauves-souris » pour souligner leurs tailles démesurées).
Chez l’Oreillard, Plecotus auritus, qui vit en Europe et en Asie, la longueur des oreilles excède celle du corps et de la tête pris ensemble. De nombreuses espèces ont des oreilles si grandes que leurs bords internes se rejoignent presque; quelques-unes ont même les oreilles soudées. Quelle que soit leur forme, les pavillons auriculaires des chauves-souris sont des cornets acoustiques extrêmement efficaces. Les Vespertillionidae et d’autres familles possèdent un tragus très large et volumineux qui ressemble alors à un deuxième pavillon situé à l’intérieur du pavillon principal (dans l’oreille humaine, le tragus est le petit appendice charnu qui se trouve juste devant le conduit auditif). Le tragus est de forme variable, tantôt c’est une pointe lancéolée, tantôt un bouton arrondi ou anguleux supporté par une tige étroite, et d’une espèce à l’autre, il y a toute une gamme de détails différents. Le tragus a pour fonction essentielle de concentrer le faisceau d’échos reçus ou plutôt d’améliorer le diagramme de directivité, et par conséquent la réceptivité de l’oreille en éliminant les zones d’ombres sonores. Sauf une exception, les chauves-souris du groupe des Rhinolophidae n’ont pas de tragus, ou s’il existe sa taille est fort réduite; en revanche, leurs oreilles sont grandes et très mobiles. Chez elles, la concentration du faisceau d’ultrasons est réalisée à l’émission par les feuilles nasales qui entourent les narines; celles-ci se trouvent séparées par un intervalle d’une demi-longueur d’onde, au fond de l’entonnoir formé par les feuilles.
On peut donc admettre que l’écholocation au moyen de l’effet Doppler n’exige pas une modification du diagramme de directivité de l’appareil récepteur.
Ajoutons que l’oreille interne de ces chauves-souris possède une cochlée relativement très volumineuse, mais on ignore l’intérêt de ce caractère pour le système sonar; on a supposé qu’il s’agissait d’une sorte de filtre, mais la chose n’a pas été prouvée. Les chauves-souris pêcheuses des genres Noctilio (superfamille des Emballonuridae) et Pizonyx (Vespertillionidae) vivent en Amérique; elles rasent les eaux douces ou marines et capturent de petits poissons. Elles possèdent également un système sonar, mais on ignore si elles l’utilisent pour repérer leurs proies. Il est certain qu’elles ne s’en servent pas pour obtenir des échos des poissons, même si ces derniers nagent à proximité de la surface, mais il est possible que les tourbillons créés dans l’eau par ces animaux produisent un signal qui les guide. En Amérique du Sud et dans le sud de l’Amérique du Nord vivent des chauves-souris dont le nez porte des feuilles nasales moins compliquées que celles de leurs homologues de l’Ancien Monde. Les animaux forment la grande famille des Phyllostomatidae. Leur feuille nasale, de forme et de taille très variable selon l’espèce fait défaut dans quelques genres, mais toutes possèdent un tragus dans l’oreille. La plupart ont un régime végétarien et se nourrissent de pollen, de nectar et du liquide qui sort de certains fruits. Les Vampires n’ont pas de feuille nasale, mais leur museau est bizarrement plissé. Ce sont des Chiroptères hématophages : après avoir percé la peau de la victime endormie à l’aide de leurs incisives tranchantes comme une lame de rasoir, ils aspirent le sang qui s’écoule de la plaie. Le sonar de ces chauves-souris diffère profondément de celui des espèces insectivores. Il émet des impulsions dont la fréquence très élevée dépasse 100 kHz, sans modulation de fréquence et qui ont une intensité si faible qu’il est difficile de les enregistrer avec les appareils couramment employés pour étudier les ultrasons des chauves-souris insectivores. Les spécialistes du sonar des Chiroptères disent qu’elles chuchotent. Cette différence est évidemment en corrélation avec le caractère statique de leurs sources de nourriture, mais on n’a pas encore étudié en détail ce type de sonar. On dit que des chiens ne sont pas attaqués par les Vampires parce qu’ils sont capables d’entendre les ultrasons et que le sonar de ces chauves-souris les éveille.
Les Chiroptères frugivores des régions tropicales de l’Ancien Monde (les Mégachiroptères) n’utilisent pas de système sonar, sauf exception. Ils ont une excellente vision, trouvent leur nourriture à l’aide de leur odorat et, généralement, se reposent à l’air libre. Seule la chauve-souris des hypogées (Roussettus aegyptiacus) possède un sonar; les dortoirs de cette espèce occupent les recoins des cavernes, des tombeaux, et des temples, là où règne une obscurité complète. Les signaux émis sont audibles et ultrasonores, et leur fréquence propre varie entre 7 et 100 kHz; ils durent à peu près 5 millisecondes et sont répétés au rythme de 7 par seconde. Ces émissions sonores sont produites par des claquements de la langue sur le palais; elles servent à l’orientation dans l’obscurité, mais dès que la lumière est suffisante, les yeux reprennent un rôle prépondérant. Ces claquements n’ont pas de fréquence modulée et leur rythme augmente quand l’animal approche d’un obstacle. Il est certain que ces claquements de langue ne sont pas aussi précis que le sonar des Microchiroptères, ceci pour la simple raison que la chauve-souris ne peut pas contrôler la fréquence de ces claquements aussi bien qu’elle le voudrait.
On a supposé que le système sonar avait été acquis d’une manière indépendante par les Mégachiroptères et les Microchiroptères; et cette hypothèse est peut-être correcte, étant donné que le sonar s’est également développé chez des animaux très éloignés comme certains Oiseaux et les Cétacés. Il est donc possible que ces deux grands groupes de Mammifères aient évolués indépendamment. Certains indices permettent de penser que les Microchiroptères ont une origine polyphylétique et que les Phyllostomatidae ont évolué d’une façon particulière.
Le radar a été inventé pendant la guerre, et quand l’ennemi fut au courant de cette découverte, il imagina des mesures défensives dans le dessein de le rendre inefficace. De leur côté, les chercheurs essayèrent de prévoir ces parades et imaginèrent des contre-parades; bref, ce petit jeu prit d’importantes proportions. L’un des appareils en service sur les avions était un récepteur qui avertissait le pilote quand un radar terrestre l’avait repéré. Cela l’aidait à échapper au repérage en adoptant une trajectoire désordonnée. Un autre moyen de réduire le radar à l’impuissance consistait à le tromper en émettant des signaux destinés à brouiller ou couvrir les signaux qu’il recevait. Phénomène surprenant, on retrouve le même genre de choses dans la nature : depuis longtemps le sonar des chauves-souris est mis en échec par certains dispositifs. On a récemment démontré que divers papillons nocturnes ont un appareil auditif sensible aux fréquences émises par les Chiroptères, et qu’ils fuient rapidement quand ils sont détectés par le sonar de ces Mammifères. Dès qu’ils perçoivent les ultrasons, ils se mettent à voler d’une façon tout à fait capricieuse, et en dernier ressort ils font une chose que ne peuvent se permettre les avions, après avoir replié leurs ailes, ils se laissent tomber sur le sol et trouvent refuge dans la végétation. Tout récemment aussi, on a établi que d’autres Papillons nocturne émettaient eux-mêmes des ultrasons quand ils percevaient les signaux d’une chauve-souris; ils parvenaient ainsi à brouiller le sonar du prédateur et l’empêchaient de repérer avec précision la proie qu’il avait détectée.
Nous sommes tellement habitués à voir le monde qui nous entoure qu’il nous est difficile d’imaginer comment l’univers apparaît à un animal qui utilise un sonar. On peut supposer que le repérage à l’aide d’un train d’impulsions émises par un appareil de ce genre donne une impression analogue à celle que nous avons en explorant une pièce obscure avec une torche électrique clignotante et en éclairant chaque objet à tour de rôle. Quand une lampe clignote à la fréquence de 50 Hz, sa lumière nous paraît continue. Même à 33 Hz (cas des installations électriques d’autrefois) la lumière semble continue, à moins qu’un mouvement ayant un effet stroboscopique ne révèle le clignotement. En outre, quand nous regardons un objet, seule la partie de son image qui vient frapper la fovéa de la rétine est vue avec précision; les choses environnantes nous paraissent floues. Quand nous lisons les lignes du haut de cette page, nous les voyons distinctement, et, quoique nous percevions celles qui se trouvent en bas, nous sommes incapables de les lire tant que nous ne portons nos regards sur elles; autrement dit, nous ne les voyons correctement qu’au moment où leur image se forme sur notre fovéa, zone d’affinité visuelle maximale.
La méthode employée par une chauve-souris pour explorer le milieu environnant est identique; l’animal dirige le faisceau de son sonar sur un point précis, puis un autre, et simultanément il émet tout autour des ondes ultrasonores d’intensité plus faible. On peut donc imaginer une analogie avec notre vision, la cible du faisceau concentré étant comparable à la portion du champ visuel qui frappe la fovéa et les ondes éparpillées à ce que nous voyons d’une manière floue. Il est possible que dans un sonar où les fréquences sont modulées, les distances soient traduites en fréquences et que le résultat de l’interférence entre les impulsions émises et les échos indique une certaine distance.
Il convient de se rappeler aussi que le type d’organe sensoriel stimulé n’a aucune importance en la matière; ce peut être celui de la vision, de l’audition, éventuellement celui de l’odorat. L’essentiel réside dans l’image mentale élaborée par le cerveau quand les messages transmis par les nerfs venant des organes lui parviennent. Quoi qu’il en soit, il est raisonnable de supposer que l’image mentale produite dans le cerveau d’une chauve-souris qui utilise un sonar est aussi détaillée que celle nous construisons en nous servant de nos yeux. Les acrobaties des Chiroptères, qu’elles soient pour éviter un prédateur, ou pour se percher sur une minuscule saillie, sont alors aisément explicables.
Quoique nos connaissances sur le sonar des chauves-souris soient assez détaillées, l’exposé qui vient d’en être fait montre que la part de l’inconnu reste immense. Comment ces animaux font-ils pour ne pas confondre leurs signaux quand ils volent de concert ? N’y a-t-il pas d’embouteillage dans leurs sociétés ? On ne peut admettre que chacune reconnaît sa propre voix, car les fréquences ne sont probablement pas tout à fait les mêmes, mais il est difficile de savoir par quel processus s’est établie cette différenciation. Comment l’oreille de la chauve-souris, sensible au faible écho, est-elle protégée contre les signaux émis par les animaux voisins ? Enfin, comment un individu peut-il éviter que ses oreilles soient temporairement assourdies par les ultrasons qu’il émet lui-même et qui l’empêcheraient de percevoir l’écho ? Des travaux récents ont montré que les potentiels électriques produits dans le cerveau par les signaux à fréquence modulée sont tels que les effets du signal sont abolis. On retrouve la même chose dans le radar : le récepteur est provisoirement débranché au cours de l’émission, de sorte qu’il ne peut recevoir le signal émis; autrement il serait évidemment paralysé et incapable de recevoir l’écho. Toutefois, cette question reste assez obscure.
La petitesse des organes qui entrent en jeu dans le système sonar rend leur étude anatomique très ardue; on retrouve les mêmes difficultés pour la mise au point d’appareils qui en simulent le fonctionnement. Ajoutons que les espèces diffèrent par des détails d’ordre morphologique et physiologique. Toutes nos connaissances proviennent d’un très petit nombre d’espèces, c’est-à-dire une fraction de cet ordre si riche. Peut-être aurons-nous un jour suffisamment d’informations pour établir un programme destiné à un ordinateur capable de répondre à certaines questions particulièrement complexes. Les ressemblances qui existent entre le sonar des chauves-souris et le radar inventé par l’homme nous aident mieux à comprendre le premier, mais la complexité du radar n’est rien comparée aux raffinements et à la souplesse du sonar. L’homme a encore beaucoup à faire pour égaler le produit d’une évolution longue de plusieurs millions d’années.