Avant d’aborder pleinement les signaux d’écholocation et leur représentation dans le domaine fréquentiel (sous forme de sonagrammes), il est indispensable de posséder quelques bases en acoustique. J’ai côtoyé de nombreux biologistes (étudiants) curieux d’en apprendre davantage sur cet aspect de l’étude comportementale des chauves-souris. Malheureusement et sans trop l’avouer, beaucoup étaient décontenancés par l’aspect souvent hermétique des explications fournies lors des colloques ou lors des discussions entre collègues. Il est pourtant question de principes de bases, mais trop souvent pris pour acquis par les spécialistes. Il est donc primordial de se replonger dans les bases de l’acoustique afin de mieux appréhender ce qui va suivre.
Le son est comparable à une onde de choc produite lors du déplacement des molécules d’air, de la même manière que des vibrations sont produites sur la surface de l’eau lors d’un jet de pierre :

Cette onde, ou vibration peut être représentée par un tracé sinusoïdal (en gris) :
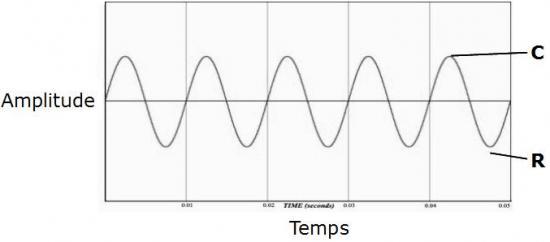
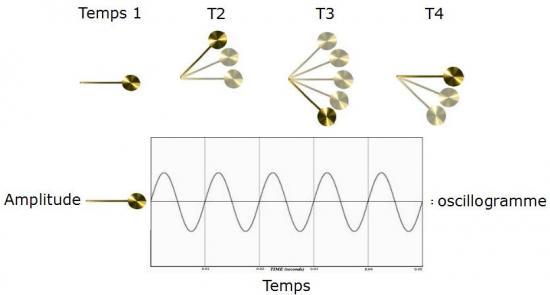
Pour caractériser le déplacement d’une onde, il suffit d’imaginer le mouvement régulier d’un pendule posé sur une feuille de papier qui avancerait au fur et à mesure pour en tracer la course :
La fréquence
Nous venons donc de comprendre ce qu’est un oscillogramme dont le signal sinusoïdal correspond à celui d’une harmonique simple (aussi appelée fréquence fondamentale). C’est à dire, d’une seule vibration dans l’air. Les sons dans la nature ont une «structure» plus complexe, car ils sont rarement constitués d’une vibration unique (Everest et Polhmann, 2009). Cet exemple simple permet tout de même d’illustrer deux autres propriétés importantes du son: la fréquence et l’amplitude. La fréquence correspond au nombre de vibrations (ou d’oscillations du pendule) par seconde :
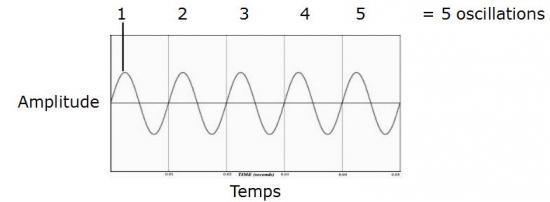
Ces cycles sont mesurés en Hertz (Hz). La voix humaine se situe en moyenne entre 100 et 400 vibrations par seconde (entre 100 et 400 Hz) avec des pics de fréquences pouvant atteindre les 5000 Hz (5 kHz) pour les chanteurs d’opéra (sopranos).
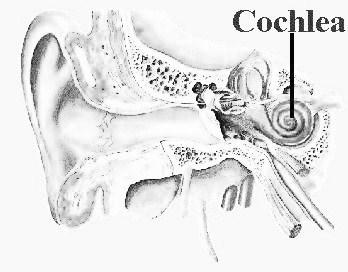
Notre appareil auditif (principalement le tympan et la cochlée) nous permet de percevoir les vibrations allant de 20 Hz à 20000 Hz (ou 20 kHz).
C’est au-dessus de ce seuil que se situe le domaine de l’ultrason. Les chauves-souris émettent des ultrasons allant de 20 kHz à 120 kHz (20000 à 120000 vibrations / sec).
L’amplitude
La deuxième propriété fondamentale du son est l’amplitude. L’amplitude représente la force (ou l’énergie) du signal sonore et elle est mesurée en décibel (dB). Elle est représentée par la hauteur atteinte par le pendule lors de chaque cycle :

L’amplitude du signal sonore est corrélée à l’énergie déployée par l’émetteur, en général, le larynx (cordes vocales) chez les mammifères (ici, coupe longitudinale d’un crâne de chauve-souris):
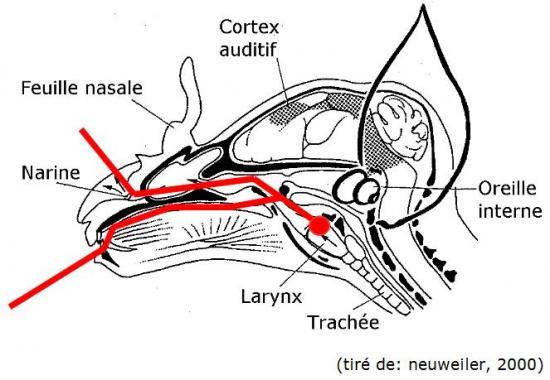
Chez les chauves-souris, les sons sont également produits par la vibration des cordes vocales à l’intérieur du larynx (point rouge sur le schéma). Ce signal voyage ensuite à travers les parties post-laryngales (pharynx, cavité nasale, bouche et lèvres ou narines et feuilles nasales, etc) où il pourra subir plusieurs transformations (notamment, de complexification par résonance), avant d’arriver jusqu’à la bouche ou les narines (en fonction des espèces), d’où il sera ensuite expulsé vers l’extérieur (Altringham, 1996; Neuweiler, 2000). Plus la quantité d’air envoyé dans la trachée est importante, plus les cordes vocales vont vibrer. Cette action augmente la force (donc l’amplitude) du signal, mais demande en contrepartie un investissement énergétique plus important (parler fort fatigue davantage; tous les enseignants vous le diront).
L’intérêt de générer un son de forte amplitude est de pouvoir le propager sur une distance relativement importante. Les signaux émis par les chauves-souris ont des pics d’amplitude compris entre 80 dB pour quelques Phyllostomidés et 120 dB chez les Rhinolophidés (Neuweiler, 2000). L’équivalent dans notre domaine de fréquences serait situé entre le bruit généré par un train à pleine vitesse (65 à 90 dB) et un avion à réaction au décollage (110 à 140 dB). Augmenter l’amplitude du signal contrebalance l’effet d’atténuation du son dans l’atmosphère (l’intensité du son est inversement proportionnelle au carré de la distance du point d’origine). Il faut également savoir que plus la fréquence d’un son est élevée plus sa propagation est limitée par la résistance de l’air. Les sons de hautes fréquences se propagent donc moins loin que les sons de basses fréquences (Altringham, 1996; Neuweiler, 2000) :
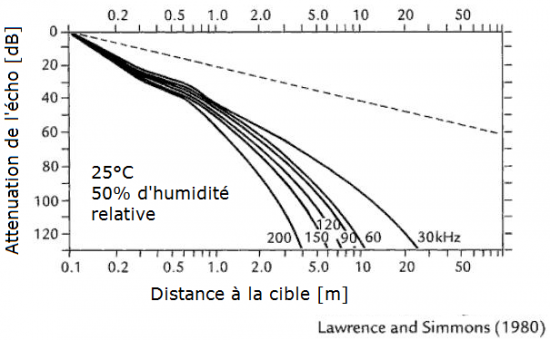
Nous le verrons par la suite (voir page suivante), cette propriété du son va avoir un impact non négligeable sur le comportement d’écholocation des chauves-souris en fonction du milieu où elles évoluent durant leur activité nocturne.
Comme mentionné précédemment, les sons dans la nature sont plus complexe, car ils sont rarement constitués d’une vibration unique. Ces tonalités supérieures («overtones» en anglais) et harmoniques sont donc d’autres propriétés inhérentes aux signaux plus complexes, c’est à dire, composés de plusieurs vibrations (Everest et Polhmann, 2009) :
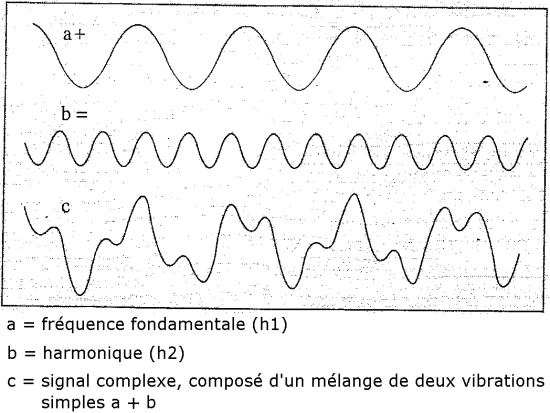
La fréquence fondamentale (également appelée première harmonique), est la fréquence la plus basse d’un signal sonore et chez les mammifères, elle est typiquement produite par les cordes vocales. En plus de cette tonalité fondamentale, un grand nombre de tonalités partielles sont produites par résonance dans le larynx et/ou lors du passage les parties post-laryngales (Neuweiler, 2000). Ces tonalités ont une fréquence systématiquement au-dessus de la fréquence fondamentale (d’où leurs noms de tonalités supérieures) et peuvent être appelées harmoniques en fonction de leur nature fréquentielle. C’est l’ensemble de ces tonalités qui donne à une voix (humaine ou non) ou à un instrument sa sonorité caractéristique. Ce sont également ces caractéristiques qui sont utilisées pour identifier les espèces (ou groupes d’espèces) à partir de leurs cris d’écholocation.
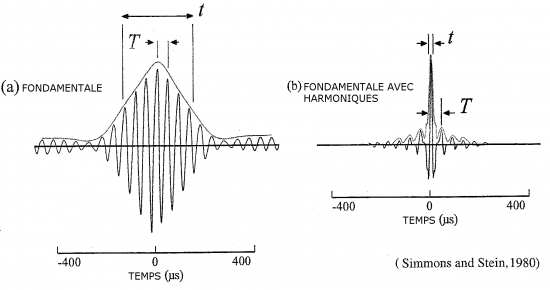
Les harmoniques sont donc en quelque sorte des copies du signal fondamental (la 1ère harmonique) à des fréquences plus élevées. Pour passer d’une harmonique à l’autre, il faut systématiquement sauter d’une octave (l’équivalent d’un saut de huit notes en musique). À chaque octave, les fréquences sont doublées. Par exemple, une fréquence fondamentale (h1) qui sera comprise entre 30 et 40 kHz, aura sa première harmonique (h2) comprise entre 60 et 80 kHz. Sa seconde harmonique (h3) sera comprise entre 90 et 120 kHz et ainsi de suite (Neuweiler, 2000). Attention toutefois, les tonalités supérieures ne sont pas forcement des harmoniques, car elles ne sont pas systématiquement produites à une octave supérieure de la fréquence fondamentale (les fréquences de ces tonalités supérieures ne sont pas décuplées). Bon nombre de biologistes se réfèrent à ces tonalités supérieures en les qualifiant à tort d’harmoniques. Le rôle des harmoniques est en quelque sorte de modeler la fréquence fondamentale pour, par exemple, augmenter l’amplitude et donc la portée du signal sonore, ou augmenter son spectre (ou bande) fréquentiel(le).
Le retrait ou l’ajout d’harmoniques dans le signal sonore permet donc aux chauves-souris d’ajuster leurs cris d’écholocation en fonction des situations auxquelles elles sont confrontées. Cette partie sera traitée plus en détail sur une autre page (à venir).
Références:
- Altringham, J. D. (1996) Bats, biology and behaviour. Oxford University Press.
- Neuweiler, G. (2000) The biology of bats. Oxford University Press.
- Everest, F. A., et Polhmann, K. C. (2009) Master handbook of acoustics, fifth edition. McGraw-Hill editions.